
이 글은 『다윈과 게놈지도가 놓친, 진화의 실체 칼슘』(윤종원) 제3장 전문입니다. 본문·수치·인용은 원고 그대로입니다.

서론: 바다를 떠난다는 것
약 3억 8천5백만 년 전, 데본기 후기의 어느 날, 일부 물고기들이 물 밖으로 기어 나왔습니다. 이것은 생명 역사에서 가장 극적인 전환점 중 하나였습니다. 38억 년 동안 생명은 오직 물속에서만 존재했습니다. 바다에서 태어나고, 바다에서 자라고, 바다에서 죽었습니다.
물은 생명의 요람이었고, 생명이 알고 있는 유일한 세계였습니다. 그런데 일부 용감한, 혹은 절박한 생물들이 이 익숙한 세계를 떠나 전혀 다른 환경으로 모험을 떠났습니다. 물에서 공기로, 아가미에서 폐로, 지느러미에서 다리로. 모든 것이 바뀌어야 했습니다. 호흡 방식이 바뀌어야 했고, 이동 방식이 바뀌어야 했고, 수분을 유지하는 방식이 바뀌어야 했습니다. 중력이라는 새로운 적과 싸워야 했습니다. 물속에서는 부력이 몸무게의 대부분을 지탱해 주었지만, 육지에서는 자신의 골격이 전체 체중을 감당해야 했습니다. 피부는 건조해지고, 눈은 공기 중에서 초점을 맞춰야 했으며, 귀는 물이 아닌 공기의 진동을 감지해야 했습니다.
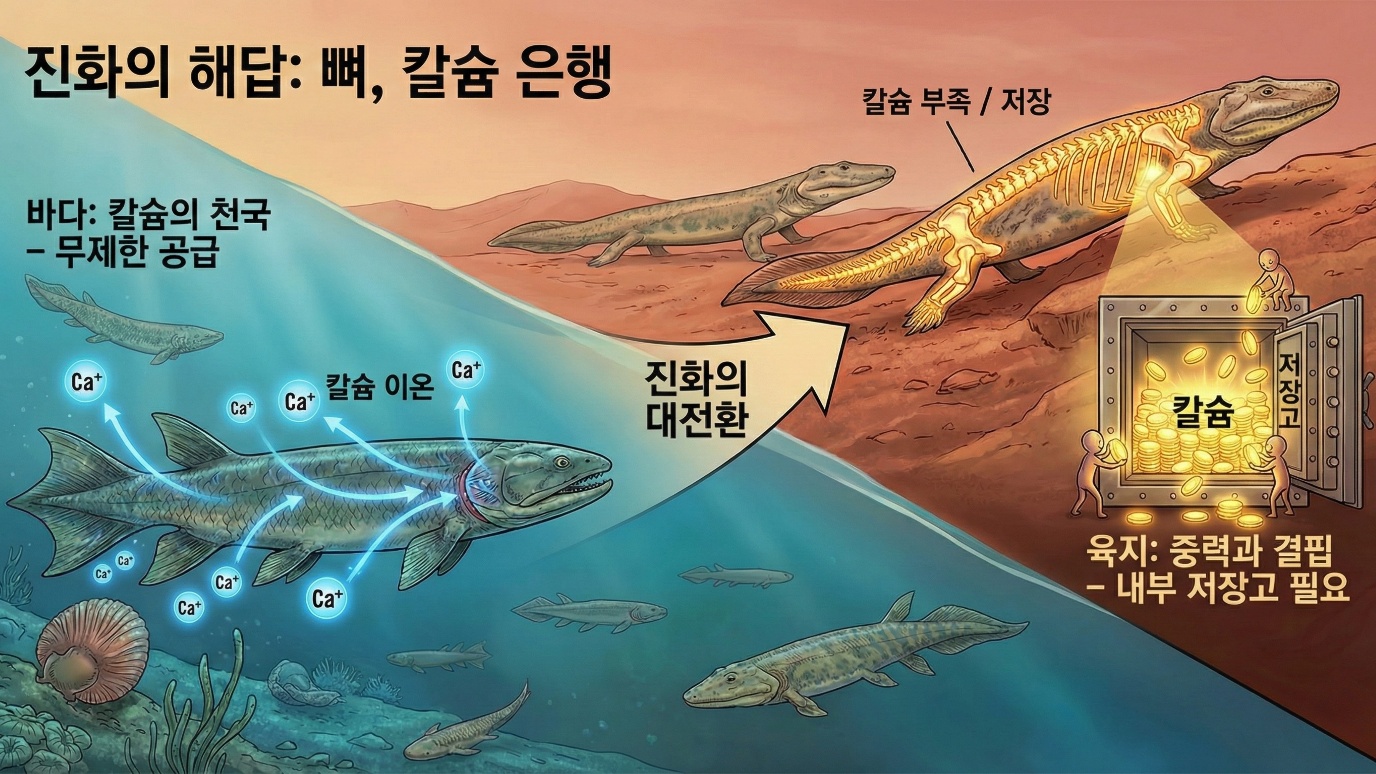
그러나 이 모든 도전 중에서 가장 간과되어 온 것이 있습니다. 바로 칼슘입니다. 우리는 육상 진출을 이야기할 때 호흡과 이동과 건조함에 대해서는 많이 이야기하지만, 칼슘에 대해서는 거의 이야기하지 않습니다. 교과서에서도 육상 진출의 핵심 혁신으로 폐와 다리와 양막을 언급하지, 칼슘 항상성 시스템을 언급하는 경우는 드뭅니다. 그러나 칼슘 문제야말로 육상 생활의 성패를 가른 결정적 요인이었습니다. 바다는 칼슘의 천국입니다. 바닷물에는 칼슘이 풍부하게 녹아 있고, 물고기는 아가미를 통해 이 칼슘을 언제든 흡수할 수 있습니다. 혈중 칼슘 농도가 조금이라도 떨어지면 아가미에서 더 흡수하면 그만입니다. 칼슘 걱정이라는 것이 존재하지 않는 세계입니다. 그런데 육지는 완전히 다릅니다. 공기 중에는 칼슘이 없습니다. 아가미도 없습니다. 오직 먹이를 통해서만 칼슘을 섭취할 수 있는데, 먹이의 칼슘 함량은 매우 가변적이고 먹이를 구하지 못하는 날도 있습니다. 칼슘이 풍부한 환경에서 칼슘이 부족한 환경으로의 이주. 이것이 육상 진출의 숨겨진 도전이었습니다.
제1장에서 우리는 칼슘이 세포의 거의 모든 기능에 관여하는 신호 분자라는 것을 배웠습니다. 근육이 수축하려면 칼슘이 필요하고, 신경이 신호를 전달하려면 칼슘이 필요하고, 심장이 박동하려면 칼슘이 필요합니다. 칼슘 없이는 단 한 순간도 살 수 없습니다. 그런데 육지에서는 이 필수 원소의 공급이 불안정합니다. 어떻게 해야 할까요? 진화가 찾아낸 답은 내부 저장고와 정밀한 조절 시스템이었습니다. 뼈를 거대한 칼슘 은행으로 만들고, 호르몬 시스템으로 이 은행의 입출금을 관리하는 것입니다. 칼슘이 풍부할 때 뼈에 저장해 두었다가, 칼슘이 부족할 때 뼈에서 꺼내 쓰는 것입니다. 이것이 바로 칼슘 항상성 시스템이며, 이 시스템의 진화가 육상 생활을 가능하게 만들었습니다. 이 장에서는 이 시스템이 어떻게 진화했는지, 왜 진화해야 했는지, 그리고 이 시스템이 칼슘 제어 능력의 진화에서 어떤 위치를 차지하는지 살펴보겠습니다.
두 세계의 차이: 칼슘 환경의 격변
바다와 육지의 칼슘 환경이 얼마나 다른지 구체적인 숫자로 살펴봅시다. 현대 바닷물의 칼슘 농도는 리터당 약 400에서 420밀리그램입니다. 이 농도에서 해수어는 아가미를 통해 칼슘을 흡수할 수 있습니다. 아가미는 호흡 기관이면서 동시에 이온 교환 기관이기도 합니다. 아가미의 상피세포에는 칼슘 통로와 칼슘 펌프가 있어서, 바닷물에서 칼슘을 능동적으로 흡수하거나 필요에 따라 배출할 수 있습니다. 사실 바다 물고기에게 칼슘은 부족한 것이 아니라 오히려 과잉입니다. 제2장에서 살펴본 것처럼, 캄브리아기 생물들이 껍질을 만들기 시작한 것도 칼슘 과잉에 대한 해독 반응이었을 수 있습니다. 바다에서는 칼슘이 너무 많아서 문제였지, 부족해서 문제였던 적은 거의 없었습니다. 바다는 말 그대로 칼슘의 천국이었습니다.
그런데 민물은 다릅니다. 민물의 칼슘 농도는 대개 리터당 약 10에서 50밀리그램 수준으로, 바닷물의 약 10분의 1에서 40분의 1에 불과합니다. 민물 물고기는 이미 칼슘이 부족한 환경에서 살고 있습니다. 아가미를 통해 칼슘을 흡수하기는 하지만, 바닷물에서처럼 풍족하게 흡수할 수는 없습니다. 민물 물고기의 아가미에는 바다 물고기보다 더 효율적인 칼슘 흡수 시스템이 발달해 있는데, 이것은 칼슘이 부족한 환경에 대한 적응입니다. 육상으로 진출한 최초의 척추동물들은 아마도 민물 환경을 거쳐 육지로 올라왔을 것입니다. 화석 기록에 따르면, 최초의 사지동물인 틱타알릭이나 아칸토스테가는 민물이나 기수 환경에서 살았던 것으로 추정됩니다. 이들은 이미 칼슘이 덜 풍부한 환경에 어느 정도 적응해 있었을 것입니다. 그러나 육지는 민물보다 더욱 극단적인 환경이었습니다. 육상 동물은 물에서 칼슘을 흡수할 수 없습니다. 아가미가 없기 때문입니다. 오직 먹이를 통해서만 칼슘을 섭취할 수 있는데, 먹이의 칼슘 함량은 매우 가변적입니다. 뼈째 먹는 작은 동물이나 칼슘이 풍부한 식물을 먹으면 칼슘을 많이 섭취할 수 있지만, 근육만 먹거나 칼슘이 적은 식물을 먹으면 칼슘 섭취가 부족합니다. 게다가 먹이를 아예 구하지 못하는 날도 있습니다. 사냥에 실패하거나 계절적으로 먹이가 부족할 때, 칼슘 섭취도 함께 부족해집니다. 바다에서는 상상할 수 없는 상황입니다. 바다에서는 먹이를 못 구해도 아가미를 통해 칼슘은 흡수할 수 있었습니다. 그러나 육지에서는 먹이가 없으면 칼슘도 없습니다.
칼슘 공급의 불안정성만이 문제가 아니었습니다. 칼슘 수요도 증가했습니다. 물속에서는 부력이 체중의 대부분을 지탱해 줍니다. 물고기가 물속에서 무게를 거의 느끼지 못하는 이유입니다. 그래서 물고기의 골격은 상대적으로 약하고 유연해도 괜찮습니다. 지느러미를 움직이고 방향을 조절하는 정도의 지지력만 있으면 됩니다. 그러나 육지에서는 상황이 완전히 다릅니다. 전체 체중을 골격이 지탱해야 합니다. 중력이라는 새로운 힘에 맞서야 합니다. 이것은 더 단단하고 더 강한 뼈를 요구하며, 더 단단하고 강한 뼈는 더 많은 칼슘을 필요로 합니다. 중력은 또한 골격에 지속적인 스트레스를 가합니다. 걸을 때마다, 뛸 때마다 뼈에 충격이 가해집니다. 뼈는 이 스트레스에 적응해야 합니다. 스트레스가 많이 가해지는 부위는 더 강화되어야 하고, 스트레스가 적은 부위는 칼슘을 아껴야 합니다. 이것은 뼈가 정적인 구조가 아니라 동적으로 재형성되어야 함을 의미합니다. 칼슘 공급은 불안정해지는데 칼슘 수요는 증가하는 상황. 이 문제를 해결하지 못하면 육상 생활은 불가능합니다. 근육이 수축하지 못하고, 신경이 신호를 보내지 못하고, 심장이 멈춥니다. 진화는 이 문제를 어떻게 해결했을까요?
부갑상선: 아가미에서 진화한 육상 생활의 열쇠
인간의 목에는 네 개의 작은 기관이 있습니다. 갑상선 뒤쪽에 붙어 있어서 부갑상선이라고 불리는 이 기관들은 크기가 쌀알 정도에 불과합니다. 무게를 다 합쳐도 약 120밀리그램 정도밖에 안 됩니다. 워낙 작아서 19세기까지 의사들도 그 존재를 잘 몰랐습니다. 갑상선 수술을 할 때 우연히 함께 제거해 버리는 경우가 많았는데, 그러면 환자가 심한 근육 경련을 일으키며 죽는 일이 빈번했습니다. 처음에는 왜 그런지 몰랐습니다. 갑상선을 제거해서 그런 줄 알았습니다. 그러나 연구가 진행되면서 진짜 원인이 밝혀졌습니다. 갑상선이 아니라 그 뒤에 붙어 있던 작은 기관들, 즉 부갑상선이 함께 제거되었기 때문이었습니다. 부갑상선 없이는 혈중 칼슘 농도가 급격히 떨어지고, 칼슘이 부족해진 근육이 통제 불능의 경련을 일으켜 죽음에 이르렀던 것입니다. 이토록 작은 기관이 생명을 좌우하는 열쇠였습니다.
부갑상선은 부갑상선호르몬을 분비합니다. 영어 약자로 PTH라고 불리는 이 호르몬의 주된 역할은 혈중 칼슘 농도를 높이는 것입니다. 어떻게 작동할까요? 부갑상선에는 칼슘감지수용체라는 특별한 단백질이 있습니다. 이 수용체는 혈중 칼슘 농도를 실시간으로 모니터링합니다. 마치 온도조절기가 실내 온도를 감지하여 난방을 조절하듯이, 칼슘감지수용체는 혈중 칼슘 농도를 감지하여 호르몬 분비를 조절합니다. 칼슘 이온이 수용체에 결합하면 부갑상선호르몬 분비가 억제되고, 칼슘 이온이 수용체에서 떨어지면 부갑상선호르몬 분비가 증가합니다. 이것은 매우 정교한 되먹임 시스템입니다. 혈중 칼슘 농도가 조금이라도 떨어지면 부갑상선이 즉시 감지하여 호르몬을 분비하고, 호르몬이 칼슘 농도를 높이면 분비가 줄어듭니다. 이 시스템 덕분에 혈중 칼슘 농도는 데시리터당 약 8.5에서 10.5밀리그램이라는 좁은 범위 안에서 유지됩니다.
부갑상선호르몬은 세 가지 방법으로 혈중 칼슘 농도를 높입니다. 가장 빠르고 직접적인 방법은 뼈에서 칼슘을 방출시키는 것입니다. 부갑상선호르몬은 뼈세포와 뼈흡수세포에 작용하여 뼈에 저장된 칼슘을 혈액으로 내보냅니다. 뼈는 칼슘의 거대한 저장고이고, 부갑상선호르몬은 이 저장고의 문을 여는 열쇠입니다. 두 번째 방법은 신장에서 칼슘 재흡수를 촉진하는 것입니다. 신장은 혈액을 여과하여 노폐물을 소변으로 배출하는데, 이 과정에서 칼슘도 일부 여과됩니다. 그러나 칼슘은 소중한 자원이므로 다시 혈액으로 끌어들여야 합니다. 부갑상선호르몬은 신장의 세뇨관에서 칼슘 재흡수를 증가시켜, 소변으로 빠져나가려던 칼슘을 다시 혈액으로 되돌립니다. 세 번째 방법은 신장에서 비타민D 활성화를 촉진하는 것입니다. 비타민D는 장에서 칼슘 흡수를 증가시키는 호르몬인데, 부갑상선호르몬이 비타민D의 활성화를 촉진하여 간접적으로 칼슘 흡수를 높입니다.
부갑상선은 어디서 왔을까요? 2004년, 학술지 PNAS에 발표된 킹스칼리지 런던의 앤서니 그레이엄 연구팀의 논문은 놀라운 답을 제시했습니다. 부갑상선은 물고기의 아가미에서 진화했다는 것입니다. 그레이엄 연구팀은 Gcm-2라는 유전자에 주목했습니다. 이 유전자는 포유류에서 부갑상선 발달에 필수적입니다. Gcm-2 유전자가 없으면 부갑상선이 형성되지 않습니다. 연구팀은 물고기에도 이 유전자가 있는지 조사했습니다. 결과는 놀라웠습니다. 제브라피시와 곰상어 모두 Gcm-2 유전자를 가지고 있었고, 이 유전자는 아가미에서 발현되었습니다. 물고기의 아가미는 호흡 기관일 뿐만 아니라 칼슘 흡수에도 중요한 역할을 합니다. 아가미에는 칼슘감지수용체도 있고, 부갑상선호르몬과 유사한 펩타이드도 발현됩니다. 기능적으로 아가미는 이미 부갑상선과 비슷한 역할을 하고 있었던 것입니다. 물고기가 육상으로 올라올 때 아가미는 호흡 기능을 잃었지만, 칼슘 조절 기능은 새로운 기관으로 이전되었습니다. 아가미 조직의 일부가 목 안쪽으로 이동하여 독립된 기관, 즉 부갑상선이 된 것입니다. 진화는 기존 구조를 새로운 목적에 맞게 재활용한 것입니다.
독립된 기관으로서의 부갑상선은 양서류에서 처음 출현합니다. 개구리에는 명확한 부갑상선이 있고, 이 부갑상선은 부갑상선호르몬을 분비하며 혈중 칼슘 농도 조절에 관여합니다. 양서류의 부갑상선은 네 번째 인두낭에서 발달하는데, 이것은 물고기에서 아가미가 발달하는 위치와 정확히 같습니다. 발생학적으로 부갑상선과 아가미가 같은 조직에서 유래했다는 결정적 증거입니다. 왜 양서류에서 부갑상선이 필요했을까요? 양서류는 물과 육지를 오가는 생활을 합니다. 올챙이일 때는 물속에서 아가미로 호흡하고 칼슘도 아가미로 흡수할 수 있지만, 변태하여 성체가 된 후에는 상당 시간을 육지에서 보냅니다. 물에 있을 때는 피부를 통해 어느 정도 칼슘을 흡수할 수 있지만, 육지에 있을 때는 이 방법을 쓸 수 없습니다. 따라서 체내 칼슘 저장고인 뼈를 효율적으로 관리하는 시스템이 필요했습니다. 부갑상선은 이 필요를 충족시키기 위해 진화한 기관입니다. 양서류를 포함한 모든 육상 척추동물, 즉 사지동물은 부갑상선을 가지고 있습니다. 파충류도, 조류도, 포유류도 모두 부갑상선이 있습니다. 이것은 육상 생활에 부갑상선이 필수적이었음을 보여줍니다.
비타민D: 자외선과 칼슘의 연결
칼슘 항상성 시스템이 완성되려면 부갑상선호르몬 외에 또 하나의 요소가 필요했습니다. 바로 비타민D입니다. 비타민D의 역사는 생명의 역사만큼 오래되었지만, 칼슘 대사에서 핵심적 역할을 하게 된 것은 육상 척추동물에 이르러서입니다. 여러 진화생물학 연구기관에 따르면, 비타민D 수용체는 원래 해독과 면역 조절에 관여했을 가능성이 크며, 칼슘과 뼈 대사 조절 기능은 육상 진출 이후 그 위에 덧붙여진 것으로 보입니다. 비타민D가 칼슘 항상성에 필수적인 역할을 하게 된 것은 양서류부터이며, 비타민D가 결핍된 양서류, 파충류, 조류, 포유류는 모두 구루병이 발생합니다. 뼈가 제대로 광물화되지 않아 약하고 휘어지는 이 질환은 비타민D가 육상 동물에서 칼슘 흡수와 뼈 형성에 필수적임을 보여줍니다.
부갑상선호르몬과 비타민D는 긴밀하게 협력하여 혈중 칼슘 농도를 조절합니다. 혈중 칼슘이 떨어지면 부갑상선에서 호르몬 분비가 증가하고, 이 호르몬은 신장에서 비타민D를 활성 형태로 전환시킵니다. 활성화된 비타민D는 장에서 칼슘 흡수를 촉진합니다. 동시에 부갑상선호르몬과 활성 비타민D는 함께 뼈에서 칼슘 방출을 촉진합니다. 반대로 혈중 칼슘이 높아지면 부갑상선호르몬 분비가 감소하고, 비타민D 활성화도 줄어들어 장에서 칼슘 흡수가 감소하고 뼈에서 칼슘 방출도 줄어듭니다. 이 정교한 되먹임 고리가 혈중 칼슘 농도를 좁은 범위 안에서 유지합니다. 육상 척추동물이 칼슘 항상성에 비타민D를 활용하게 된 것은 진화적으로 합리적인 선택이었습니다. 육상 동물은 바다 동물보다 훨씬 많은 자외선에 노출되고, 피부에서 자외선에 의해 비타민D가 합성되는 것은 자연스러운 일입니다. 자외선 노출이라는 환경 신호를 칼슘 대사와 연결시킨 것은 우아한 해결책이었습니다.
뼈: 살아있는 칼슘 저장고
많은 사람들이 뼈를 정적인 구조물로 생각합니다. 일단 형성되면 변하지 않는, 마치 집의 기둥 같은 존재라고 생각합니다. 그러나 이것은 큰 오해입니다. 뼈는 살아있는 조직입니다. 뼈 안에는 혈관이 흐르고, 신경이 지나가고, 다양한 세포들이 활동합니다. 그리고 뼈는 끊임없이 분해되고 재형성됩니다. 성인의 골격은 약 10년에 한 번씩 완전히 교체됩니다. 지금 당신의 뼈를 이루고 있는 칼슘 원자들은 10년 전과 완전히 다른 원자들입니다. 이 과정을 뼈 재형성이라고 합니다. 뼈흡수세포가 오래된 뼈를 분해하고, 뼈형성세포가 새로운 뼈를 만듭니다. 정상적인 상황에서 이 두 과정은 균형을 이룹니다. 분해되는 양만큼 새로 형성되므로 전체 뼈의 양은 일정하게 유지됩니다. 왜 이렇게 복잡한 시스템이 필요할까요? 두 가지 이유가 있습니다.
첫째, 기계적 적응입니다. 뼈는 스트레스를 받는 부위가 강화되어야 합니다. 운동선수의 뼈가 일반인보다 강한 이유입니다. 테니스 선수의 라켓을 잡는 팔뼈가 반대쪽 팔뼈보다 더 굵고 강합니다. 발레리나의 발뼈는 일반인보다 훨씬 치밀합니다. 뼈 재형성을 통해 스트레스 분포에 따라 뼈 구조를 최적화할 수 있습니다. 스트레스가 많이 가해지는 부위에서는 뼈형성이 뼈흡수보다 많이 일어나 뼈가 강화되고, 스트레스가 적은 부위에서는 뼈흡수가 뼈형성보다 많이 일어나 칼슘이 다른 곳으로 재배치됩니다. 이것은 한정된 칼슘 자원을 가장 효율적으로 사용하는 방법입니다. 둘째, 칼슘 저장고 기능입니다. 뼈는 체내 칼슘의 99퍼센트 이상을 저장하고 있습니다. 필요할 때 이 칼슘을 꺼내 쓰려면 뼈를 분해해야 합니다. 뼈 재형성 시스템이 없다면 이것이 불가능합니다. 뼈가 한번 형성되면 영구적으로 고정되는 구조라면, 혈중 칼슘이 부족할 때 뼈에서 칼슘을 꺼낼 방법이 없습니다. 뼈 재형성 시스템은 뼈를 동적인 칼슘 저장고로 만들어, 필요에 따라 칼슘을 저장하고 인출할 수 있게 합니다.
물고기도 뼈가 있습니다. 경골어류는 이름 그대로 뼈를 가지고 있습니다. 그런데 물고기 뼈와 육상 동물 뼈는 중요한 차이가 있습니다. 많은 경골어류에서 뼈 재형성은 포유류처럼 전신 칼슘 항상성의 중심 축이라기보다는, 골격 지지와 성장에 초점을 둔 형태로 이루어집니다. 물고기는 아가미를 통해 바닷물에서 칼슘을 직접 흡수할 수 있기 때문에, 육상 척추동물만큼 뼈를 칼슘 은행으로 의존하지는 않습니다. 칼슘이 필요하면 아가미에서 더 흡수하면 됩니다. 굳이 뼈를 분해할 필요가 없습니다. 그러나 양서류부터 상황이 달라집니다. 양서류의 뼈에는 뼈흡수세포가 있고, 활발한 뼈 재형성이 일어납니다. 부갑상선호르몬과 활성 비타민D가 뼈흡수세포의 형성과 기능을 조절합니다. 뼈가 진정한 칼슘 저장고로 기능하기 시작한 것입니다. 파충류, 조류, 포유류로 갈수록 뼈 재형성 시스템은 더욱 정교해집니다. 포유류의 뼈에는 뼈세포가 풍부하게 분포합니다. 뼈세포는 기계적 스트레스를 감지할 뿐만 아니라, 신장이나 췌장 같은 다른 장기들과 호르몬으로 소통하는 내분비 기능도 수행합니다. 뼈는 침묵하는 기둥이 아니라, 온몸과 끊임없이 대화하는 거대한 내분비 기관이기도 합니다.
인체에서 칼슘이 어떻게 분포되어 있는지 살펴봅시다. 성인의 체내에는 약 1에서 1.2킬로그램의 칼슘이 있습니다. 이 중 99퍼센트는 뼈와 치아에 있습니다. 인산칼슘, 주로 하이드록시아파타이트 형태로 저장되어 있습니다. 나머지 1퍼센트는 혈액과 세포에 있습니다. 이 1퍼센트가 생리적으로 중요합니다. 혈중 칼슘은 근육 수축, 신경 전달, 혈액 응고, 호르몬 분비 등 거의 모든 생리 기능에 관여합니다. 1퍼센트라고 무시할 수 없습니다. 절대량으로 따지면 약 10에서 12그램입니다. 그리고 이 양은 매우 좁은 범위, 데시리터당 약 8.5에서 10.5밀리그램 내에서 유지되어야 합니다. 이 범위를 벗어나면 심각한 문제가 발생합니다. 칼슘이 너무 낮으면 근육이 통제 불능으로 수축하는 테타니가 일어나고, 칼슘이 너무 높으면 심장 부정맥과 의식 장애가 발생합니다. 뼈의 99퍼센트는 이 1퍼센트를 위한 저장고입니다. 혈중 칼슘이 떨어지면 뼈에서 꺼내 쓰고, 충분하면 다시 저장합니다. 뼈는 구조적 지지대이기도 하지만, 동시에 거대한 칼슘 은행이기도 합니다.
한 가지 더 주목할 점이 있습니다. 뼈의 주성분인 하이드록시아파타이트에는 칼슘뿐만 아니라 인도 저장되어 있습니다. 인은 에너지 대사의 핵심인 ATP의 필수 구성 요소이자 DNA와 RNA의 골격을 이루는 원소입니다. 제1장에서 우리는 칼슘이 ATP의 인산기와 결합하여 침전물을 형성할 수 있다는 것을 배웠습니다. 흥미롭게도, 뼈에서는 바로 이 반응이 의도적으로 일어납니다. 칼슘과 인산이 결합하여 하이드록시아파타이트라는 안정한 광물을 형성하고, 이것이 뼈의 강도를 제공합니다. 세포 안에서는 재앙인 반응이 세포 밖 뼈에서는 유용한 반응이 되는 것입니다. 육상 진출은 칼슘과 인이라는 두 가지 핵심 자원을 뼈라는 하나의 금고에 통합 관리하게 된 사건이기도 합니다.
진화적 통합: 칼슘 제어의 세 번째 차원
이제 칼슘 제어 능력의 진화를 정리해 봅시다. 제1장에서 우리는 첫 번째 차원을 살펴보았습니다. 세포 내 칼슘 제어입니다. 모든 세포는 세포질 칼슘 농도를 약 100나노몰로 유지합니다. 세포 밖과 1만 배 이상의 농도 차이를 유지하면서, 필요할 때만 칼슘을 일시적으로 유입시켜 신호로 사용합니다. 이것은 38억 년 전부터 보존된 가장 근본적인 칼슘 제어입니다. 대장균에서 인간 신경세포까지, 모든 생명체가 공유하는 능력입니다. 제2장에서 우리는 두 번째 차원을 살펴보았습니다. 세포 외 칼슘 구조물입니다. 캄브리아기 대폭발에서 동물들은 세포 밖에 칼슘 기반 껍질과 골격을 만들기 시작했습니다. 세포 내 칼슘 관리 능력이 세포 밖으로 확장된 것입니다. 위험한 독을 밖으로 밀어내던 시스템이, 그 독을 단단한 갑옷으로 바꾸는 시스템으로 진화했습니다. 이 장에서 우리는 세 번째 차원을 살펴보았습니다. 전신 칼슘 항상성입니다. 육상 척추동물에서 진화한 부갑상선호르몬과 비타민D 시스템입니다. 이 시스템은 혈중 칼슘 농도를 조절하고, 뼈라는 내부 저장고를 관리합니다. 각 차원은 이전 차원 위에 구축되었습니다. 세포 내 칼슘 제어가 없었다면 세포 외 구조물을 만들 수 없었을 것입니다. 칼슘을 정밀하게 다루는 능력이 있어야 원하는 위치에 원하는 형태로 껍질과 골격을 형성할 수 있기 때문입니다. 세포 외 구조물, 즉 뼈가 없었다면 전신 칼슘 항상성 시스템이 작동할 저장고가 없었을 것입니다.
진화의 특징은 완전히 새로운 것을 만들기보다 기존 것을 재활용한다는 점입니다. 칼슘 항상성 시스템도 예외가 아닙니다. 부갑상선호르몬 유전자는 물고기에도 존재합니다. 그러나 어류에서 이 펩타이드는 여러 조직에서 분산된 형태로 작동하며 국소적 작용 비중이 큽니다. 사지동물에서처럼 부갑상선에서 혈류로, 혈류에서 전신으로라는 명확한 내분비 축으로 정리되어 있지는 않습니다. 양서류 이후 부갑상선이 출현하면서 이 펩타이드가 오늘날 우리가 아는 전신 칼슘 호르몬으로 재편되었다는 것이 최근 진화생리학의 해석입니다. 비타민D 수용체도 물고기에 있습니다. 그러나 물고기에서 이 수용체의 역할은 주로 해독과 면역 조절입니다. 칼슘 항상성 조절은 부차적입니다. 육상 척추동물에서 비타민D 수용체는 칼슘 항상성의 핵심 조절자가 되었습니다. 부갑상선 자체도 물고기 아가미에서 유래했습니다. 칼슘 흡수 기능을 하던 아가미 조직이 육상으로 올라오면서 독립 기관으로 전환된 것입니다. 기존 유전자, 기존 단백질, 기존 조직이 새로운 맥락에서 새로운 기능을 획득했습니다. 이것이 진화의 방식입니다.
혈중 칼슘 농도는 왜 이렇게 정밀하게 조절되어야 할까요? 앞서 제1장에서 살펴본 것처럼, 칼슘은 세포의 거의 모든 기능에 관여하는 신호 분자입니다. 근육 수축, 신경 전달, 호르몬 분비, 세포 분열. 이 모든 과정이 칼슘 신호에 의존합니다. 이 신호가 정상적으로 작동하려면 기저 칼슘 농도가 일정해야 합니다. 기저 농도가 흔들리면 신호 대 잡음비가 떨어집니다. 신호가 왔는지 안 왔는지 구분이 어려워집니다. 세포질 칼슘 농도가 100나노몰로 유지되어야 하듯이, 세포 밖 칼슘 농도도 일정하게 유지되어야 합니다. 세포 밖 칼슘 농도가 일정해야 세포 안으로 유입되는 칼슘의 양을 예측할 수 있고, 칼슘 신호가 정상적으로 작동합니다. 혈중 칼슘 농도는 세포 외 칼슘 농도를 결정합니다. 결론적으로, 전신 칼슘 항상성 시스템은 세포 수준의 칼슘 신호전달이 정상적으로 작동하도록 환경을 조성하는 역할을 합니다. 거시적 시스템이 미시적 과정을 지원하는 것입니다.
결론: 바다를 몸 안에 품다
약 3억 8천5백만 년 전 육상 진출은 척추동물 진화의 결정적 전환점이었습니다. 중력에 대항하여 몸을 지탱하고, 공기로 호흡하고, 건조한 환경에서 수분을 유지해야 했습니다. 그러나 가장 근본적인 도전 중 하나는 칼슘이었습니다. 바다라는 칼슘 천국을 떠나 칼슘 사막으로 들어간 것입니다. 아가미라는 칼슘 흡수 기관도 잃었습니다. 새로운 해결책이 필요했습니다. 진화의 답은 내부 저장고와 정밀한 조절 시스템이었습니다. 뼈는 단순한 지지 구조에서 거대한 칼슘 은행으로 기능이 확장되었습니다. 부갑상선과 비타민D로 구성된 호르몬 시스템이 이 은행을 관리하기 시작했습니다.
이 시스템은 완전히 새로 만들어진 것이 아닙니다. 물고기에 이미 존재하던 유전자, 단백질, 조직이 새로운 맥락에서 새로운 기능을 획득한 것입니다. 아가미에서 부갑상선으로, 국소 인자에서 전신 호르몬으로. 진화는 기존 재료를 재활용하여 새로운 도전에 대응했습니다. 제2장에서 우리는 척추동물이 뼈라는 형태의 휴대용 고체 바다를 몸속에 챙겨서 육지로 올라왔다고 했습니다. 이 표현은 비유가 아닙니다. 뼈에 저장된 칼슘과 인, 그리고 이것을 관리하는 호르몬 시스템은 말 그대로 바다의 이온 환경을 몸 안에 재현한 것입니다. 물고기는 바다에서 살지만, 육상 척추동물은 바다를 몸 안에 품고 삽니다. 혈액의 이온 조성이 고대 바닷물과 비슷하다는 사실은 우연이 아닙니다. 우리는 바다를 떠났지만, 바다는 우리 안에 남아 있습니다.
칼슘 제어 능력의 진화는 세 번째 차원에 도달했습니다. 세포 내 칼슘 관리에서 세포 외 구조물 형성으로, 그리고 전신 칼슘 항상성으로. 각 단계는 이전 단계 위에 구축되었습니다. 이 책의 제1부에서 우리는 칼슘 제어 능력이 어떻게 진화해 왔는지, 그리고 그것이 생명의 주요 전환점마다 어떤 역할을 했는지 살펴보았습니다. 다음 장에서는 제1부의 마지막으로, 칼슘과 고등 지능의 관계를 살펴볼 것입니다. 뇌라는 기관이 왜 그토록 칼슘에 의존하는지, 그리고 칼슘 신호전달의 정교함이 어떻게 고등 인지 기능을 가능하게 했는지 알아보겠습니다. 그리고 제2부에서는 이 정교한 시스템이 어떻게 고장 나는지 살펴볼 것입니다. 칼슘 항상성의 붕괴가 만성질환의 원인이 된다는 것이 이 책의 핵심 주장인 DIAH 가설입니다.
참고문헌
1. Okabe, M., & Graham, A. (2004). The origin of the parathyroid gland. Proceedings of the National Academy of Sciences, 101(51), 17716–17719. doi:10.1073/pnas.0406116101
2. Bouillon, R., & Suda, T. (2014). Vitamin D: calcium and bone homeostasis during evolution. BoneKEy Reports, 3, 480. doi:10.1038/bonekey.2013.214
3. Suarez-Bregua, P., Cal, L., Cañestro, C., & Rotllant, J. (2017). PTH reloaded: A new evolutionary perspective. Frontiers in Physiology, 8, 776. doi:10.3389/fphys.2017.00776
4. On, J. S., Chow, B. K., & Lee, L. T. (2015). Evolution of parathyroid hormone receptor family and their ligands in vertebrates. Frontiers in Endocrinology, 6, 28. doi:10.3389/fendo.2015.00028
5. Talmage, R. V., & Mobley, H. T. (2008). Calcium homeostasis: Reassessment of the actions of parathyroid hormone. General and Comparative Endocrinology, 156(1), 1–8. doi:10.1016/j.ygcen.2007.11.003
6. Carlberg, C. (2022). Vitamin D in the context of evolution. Nutrients, 14(15), 3018. doi:10.3390/nu14153018
7. Guerreiro, P. M., Renfro, J. L., Power, D. M., & Canario, A. V. (2007). The parathyroid hormone family of peptides: Structure, tissue distribution, regulation, and potential functional roles in calcium and phosphate balance in fish. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology, 292(2), R679–R696. doi:10.1152/ajpregu.00480.2006
8. Holick, M. F. (2018). Vitamin D: part I; from plankton and calcified skeletons (500 million years ago) to rickets. International Orthopaedics, 42(3), 475–480. doi:10.1007/s00264-018-3857-3